糖酵解 三羧酸循环最全总结

在高等植物中存在着多条呼吸代谢的生化途径,这是植物在长期进化过程中,对多变环境条件适应的体现。在缺氧条件下进行酒精发酵和乳酸发酵,在有氧条件下进行三羧酸循环和戊糖磷酸途径,还有脂肪酸氧化分解的乙醛酸循环以及乙醇酸氧化途径等(图5-2)。
图5-2 植物体内主要呼吸代谢途径相互关系示意图
一、糖酵解
己糖在细胞质中分解成丙酮酸的过程,称为糖酵解(glycolysis)。整个糖酵解化学过程于1940年得到阐明。为纪念在研究这一途径中有突出贡献的三位生物化学家:G.Embden,O.Meyerhof和J.K.Parnas,又把糖酵解途径称为EmbdenMeyerhofParnas途径,简称EMP途径(EMP pathway)。糖酵解普遍存在于动物、植物、微生物的细胞中。
(一)糖酵解的化学历程
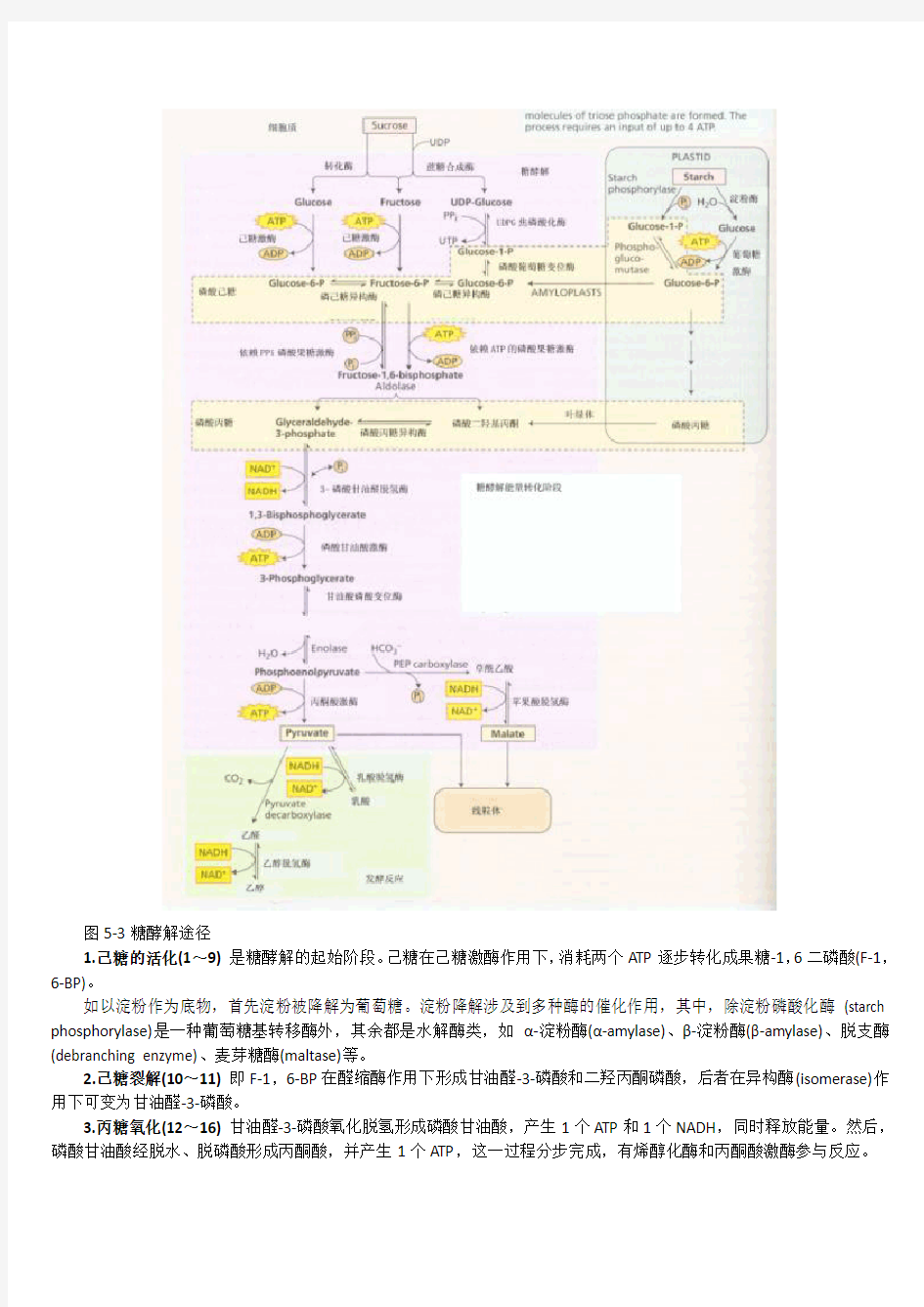
糖酵解途径(图5-3)可分为下列几个阶段:
图5-3糖酵解途径
1.己糖的活化(1~9)是糖酵解的起始阶段。己糖在己糖激酶作用下,消耗两个ATP逐步转化成果糖-1,6二磷酸(F-1,6-BP)。
如以淀粉作为底物,首先淀粉被降解为葡萄糖。淀粉降解涉及到多种酶的催化作用,其中,除淀粉磷酸化酶(starch phosphorylase)是一种葡萄糖基转移酶外,其余都是水解酶类,如α-淀粉酶(α-amylase)、β-淀粉酶(β-amylase)、脱支酶(debranching enzyme)、麦芽糖酶(maltase)等。
2.己糖裂解(10~11)即F-1,6-BP在醛缩酶作用下形成甘油醛-3-磷酸和二羟丙酮磷酸,后者在异构酶(isomerase)作用下可变为甘油醛-3-磷酸。
3.丙糖氧化(12~16)甘油醛-3-磷酸氧化脱氢形成磷酸甘油酸,产生1个ATP和1个NADH,同时释放能量。然后,磷酸甘油酸经脱水、脱磷酸形成丙酮酸,并产生1个ATP,这一过程分步完成,有烯醇化酶和丙酮酸激酶参与反应。
糖酵解过程中糖的氧化分解是在没有分子氧的参与下进行的,其氧化作用所需要的氧来自水分子和被氧化的糖分子。
在糖酵解过程中,每1mol葡萄糖产生2mol丙酮酸时,净产生2molATP和2molNADH+H+。
根据图5-3,糖酵解的总反应可归纳为:
C6H12O6+2NAD++2ADP+2H3PO4→2CH3COCOOH+2NADH+2H++2ATP (5-4)
(二)糖酵解的生理意义
1.糖酵解普遍存在于生物体中,是有氧呼吸和无氧呼吸途径的共同部分。
2.糖酵解的产物丙酮酸的化学性质十分活跃,可以通过各种代谢途径,生成不同的物质(图5-4)。
图5-4 丙酮酸在呼吸和物质转化中的作用
3.通过糖酵解,生物体可获得生命活动所需的部分能量。对于厌氧生物来说,糖酵解是糖分解和获取能量的主要方式。
4. 糖酵解途径中,除了由己糖激酶、磷酸果糖激酶、丙酮酸激酶等所催化的反应以外,多数反应均可逆转,这就为糖异生作用提供了基本途径。
二、发酵作用
生物体中重要的发酵作用有酒精发酵和乳酸发酵。在酒精发酵(alcohol fermentation)过程中,糖类经过糖酵解生成丙酮酸。然后,丙酮酸先在丙酮酸脱羧酶(pyruvic acid decarboxylase)作用下脱羧生成乙醛。
CH3COCOOH→CO2+CH3CHO (5-5)
乙醛再在乙醇脱氢酶(alcohol dehydrogenase)的作用下,被还原为乙醇。
CH3CHO+NADH+H+→CH3CH2OH+NAD+ (5-6)
在缺少丙酮酸脱羧酶而含有乳酸脱氢酶(lactic acid dehydrogenase)的组织里,丙酮酸便被NADH还原为乳酸,即乳酸发酵(lactate fermentation)。
CH3COCOOH+NADH+H+→CH3CHOHCOOH+NAD+ (5-7)
在无氧条件下,通过酒精发酵或乳酸发酵,实现了NAD+的再生,这就使糖酵解得以继续进行。
无氧呼吸过程中形成乙醇或乳酸所需的NADH+H+,一般来自于糖酵解。因此,当植物进行无氧呼吸时,糖酵解过程中形成的2分子NADH+H+就会被消耗掉(图5-5),这样每分子葡萄糖在发酵时,只净生成2分子A TP,葡萄糖中的大部分能量仍保存在乳酸或乙醇分子中。可见,发酵作用能量利用效率低,有机物耗损大,依赖无氧呼吸不可能长期维持细胞的生命活动,而且发酵产物的产生和累积,对细胞原生质有毒害作用。如酒精累积过多,会破坏细胞的膜结构;若酸性的发酵产物累积量超过细胞本身的缓冲能力,也会引起细胞酸中毒。
图5-5 NAD+与NADH的周转与丙酮酸还原之间的关系
三、三羧酸循环
糖酵解的最终产物丙酮酸,在有氧条件下进入线粒体,通过一个包括三羧酸和二羧酸的循环逐步脱羧脱氢,彻底氧化分解,这一过程称为三羧酸循环(tricarboxylic acid cycle,TCAC)。这个循环是英国生物化学家克雷布斯(H.Krebs)首先发现的,所以又名Krebs 循环(Krebs cycle)。1937年他提出了一个环式反应来解释鸽子胸肌内的丙酮酸是如何分解的,并把这一途径称为柠檬酸循环(citric acid cycle),因为柠檬酸是其中的一个重要中间产物。TCA循环普遍存在于动物、植物、微生物细胞中,是在线粒体基质中进行的。TCA循环的起始底物乙酰CoA不仅是糖代谢的中间产物,也是脂肪酸和某些氨基酸的代谢产物。因此,TCA循环是糖、脂肪、蛋白质三大类物质的共同氧化途径。
(一)三羧酸循环的化学历程
TCA循环共有9步反应(图5-6)。
1.反应(1)丙酮酸在丙酮酸脱氢酶复合体催化下氧化脱羧生成乙酰CoA,这是连结EMP与TCAC的纽带。
丙酮酸脱氢酶复合体(pyruvic acid dehydrogenase complex)是由3种酶组成的复合体,含有6种辅助因子。这3种酶是:丙酮酸脱羧酶(pyruvic acid decarboxylase)、二氢硫辛酸乙酰基转移酶(dihydrolipoyltransacetylase)、二氢硫辛酸脱氢酶(dihydrolipoic acid dehydrogenase)。6种辅助因子。6种辅助因子分别是硫胺素焦磷酸(thiamine pyrophosphate,TPP)、辅酶A (coenzyme A)、硫辛酸(lipoic acid)、FAD(flavin adenine dinucleotide)、NAD+(nicotinamide adenine dinucleotide)和Mg2+。
图5-6 三羧酸循环的反应过程
上述反应中从底物上脱下的氢是经FAD→FADH2传到NAD+再生成NADH+H+。
2.反应(2)乙酰CoA在柠檬酸合成酶催化下与草酰乙酸缩合为柠檬酸,并释放CoASH,此反应为放能反应(△G°,=-32.22kJ·mol-1)。
3.反应(3)由顺乌头酸酶催化柠檬酸脱水生成顺乌头酸,然后加水生成异柠檬酸。
4.反应(4)在异柠檬酸脱氢酶催化下,异柠檬酸脱氢生成NADH,其中间产物草酰琥珀酸是一个不稳定的β-酮酸,与酶结合即脱羧形成α-酮戊二酸。
5.反应(5)α酮戊二酸在α酮戊二酸脱氢酶复合体催化下形成琥珀酰辅酶A和NADH,并释放CO2。
α酮戊二酸脱氢酶复合体是由α酮戊二酸脱羧酶(α-ketoglutaric acid decarboxylase)、二氢硫辛酸琥珀酰基转移酶(dihydrolipoyltranssuccinylase) 及二氢硫辛酸脱氢酶所组成的,含有6种辅助因子:TPP、NAD+、辅酶A、FAD、硫辛酸及Mg2+。该反应不可逆。
6.反应(6)含有高能硫酯键的琥珀酰CoA在琥珀酸硫激酶催化下,利用硫酯键水解释放的能量,使ADP磷酸化成ATP。该反应是TCA循环中唯一的一次底物水平磷酸化,即由高能化合物水解,放出能量直接形成ATP的磷酸化作用。
7.反应(7)琥珀酸在琥珀酸脱氢酶催化下,脱氢氧化生成延胡索酸,脱下的氢生成FADH2。丙二酸、戊二酸与琥珀酸
的结构相似,是琥珀酸脱氢酶特异的竞争性抑制剂。
8.反应(8)延胡索酸经延胡索酸酶催化加水生成苹果酸。
9.反应(9)苹果酸在苹果酸脱氢酶的催化下氧化脱氢生成草酰乙酸和NADH。草酰乙酸又可重新接受进入循环的乙酰CoA,再次生成柠檬酸,开始新一轮TCA循环。
TCA循环的总反应式为:CH3COCOOH+4NAD++FAD+ADP+Pi+2H2O3CO2+4NADH+4H++FADH2+ATP(5-8)
(二)三羧酸循环的回补机制
TCA循环中某些中间产物是合成许多重要有机物的前体。例如草酰乙酸和α酮戊二酸分别是天冬氨酸和谷氨酸合成的碳架,延胡索酸是苯丙氨酸和酪氨酸合成的前体,琥珀酰CoA是卟啉环合成的碳架。如果TCA循环的中间产物大量消耗于有机物的合成,就会影响TCA循环的正常运行,因此必须有其他的途径不断地补充,这称之为TCA循环的回补机制(replenishing mechanism)。主要有三条回补途径:
1.丙酮酸的羧化丙酮酸在丙酮酸羧化酶催化下形成草酰乙酸。
Pyr+CO2+H2O+ATPOAA+ADP+Pi (5-9)
丙酮酸羧化酶的活性平时较低,当草酰乙酸不足时,由于乙酰CoA 的累积可提高该酶活性。这是动物中最重要的回补反应。
2.PEP的羧化作用在糖酵解中形成的PEP不转变为丙酮酸,而是在PEP羧化激酶作用下形成草酰乙酸,草酰乙酸再被还原为苹果酸,苹果酸经线粒体内膜上的二羧酸传递体与Pi进行电中性的交换,进入线粒体基质,可直接进入TCA 循环;苹果酸也可在苹果酸酶的作用下脱羧形成丙酮酸,再进入TCA循环都可起到补充草酰乙酸的作用。这一回补反应存在于高等植物、酵母和细菌中,动物中不存在。
PEP+CO2+H2O→OAA+Pi (5-10)
3.天冬氨酸的转氨作用天冬氨酸和α酮戊二酸在转氨酶作用下可形成草酰乙酸和谷氨酸:
ASP+α-酮戊二酸OAA+Glu (5-11)
通过以上这些回补反应,保证有适量的草酰乙酸供TCA循环的正常运转。
(三)三羧酸循环的特点和生理意义
1.在TCA循环中底物(含丙酮酸)脱下5对氢原子,其中4对氢在丙酮酸、异柠檬酸、α-酮戊二酸氧化脱羧和苹果酸氧化时用以还原NAD+,一对氢在琥珀酸氧化时用以还原FAD。生成的NADH和FADH2,经呼吸链将H+和电子传给O2生成H2O,同时偶联氧化磷酸化生成A TP。此外,由琥珀酰CoA形成琥珀酸时通过底物水平磷酸化生成ATP。因而,TCA 循环是生物体利用糖或其它物质氧化获得能量的有效途径。
2.乙酰CoA与草酰乙酸缩合形成柠檬酸,使两个碳原子进入循环。在两次脱羧反应中,两个碳原子以CO2的形式离开循环,加上丙酮酸脱羧反应中释放的CO2,这就是有氧呼吸释放CO2的来源,当外界环境中二氧化碳浓度增高时,脱羧反应减慢,呼吸作用就减弱。TCA循环中释放的CO2中的氧,不是直接来自空气中的氧,而是来自被氧化的底物和水中的氧。
3.在每次循环中消耗2分子H2O。一分子用于柠檬酸的合成,另一分子用于延胡索酸加水生成苹果酸。水的加入相当于向中间产物注入了氧原子,促进了还原性碳原子的氧化。
4.TCA循环中并没有分子氧的直接参与,但该循环必须在有氧条件下才能进行,因为只有氧的存在,才能使NAD+和FAD在线粒体中再生,否则TCA循环就会受阻。
5.该循环既是糖、脂肪、蛋白彻底氧化分解的共同途径;又可通过代谢中间产物与其他代谢途径发生联系和相互转变。
四、戊糖磷酸途径
20世纪50年代初的研究发现EMP-TCAC途径并不是高等植物中有氧呼吸的唯一途径。实验证据是,当向植物组织匀浆中添加糖酵解抑制剂(氟化物和碘代乙酸等)时,不可能完全抑制呼吸。瓦伯格(Warburg)也发现,葡萄糖氧化为磷酸丙糖可不需经过醛缩酶的反应。此后不久,便发现了戊糖磷酸途径(pentose phosphate pathway,PPP),又称己糖磷酸途径(hexose monophosphate pathway,HMP)或己糖磷酸支路(shunt)。
图5-7 戊糖磷酸途径
(一)戊糖磷酸途径的化学历程
戊糖磷酸途径是指葡萄糖在细胞质内直接氧化脱羧,并以戊糖磷酸为重要中间产物的有氧呼吸途径。该途径可分为两个阶段(图5-7)。
1.葡萄糖氧化脱羧阶段
(1)脱氢反应在葡萄糖-6-磷酸脱氢酶(glucose6phosphate dehydrogenase)的催化下以NADP+为氢受体,葡萄糖-6-磷酸(G6P)脱氢生成6-磷酸葡萄糖酸内酯(6phosphogluconolactone,6PGL)。
(2)水解反应在6-磷酸葡萄糖酸内酯酶(lactonase)的催化下,6-PGL被水解为6-磷酸葡萄糖酸(6phosphogluconate,6-PG)。反应是可逆的。
(3)脱氢脱羧反应在6-磷酸葡萄糖酸脱氢酶(6-phosphogluconate dehydrogenase)催化下,以NADP+为氢受体,6-PG氧
化脱羧,生成核酮糖-5-磷酸(Ru5P)。
本阶段的总反应是:
G6P+2NADP++H2O→Ru5P+CO2+2NADPH+2H+ (5-12)
2.分子重组阶段经过一系列糖之间的转化,最终可将6个Ru5P转变为5个G6P(图5-7中反应4~12)。
从整个戊糖磷酸途径来看,6分子的G6P经过两个阶段的运转,可以释放6分子CO2、12分子NADPH,并再生5分子G6P。戊糖磷酸途径的总反应式可写成:6G6P+12NADP++7H2O→6CO2+12NADPH+12H++5G6P+Pi (5-13)
(二)戊糖磷酸途径的特点和生理意义
1.PPP是葡萄糖直接氧化分解的生化途径,每氧化1分子葡萄糖可产生12分子的NADPH+H+,有较高的能量转化效率。
2.该途径中生成的NADPH在脂肪酸、固醇等的生物合成、非光合细胞的硝酸盐、亚硝酸盐的还原以及氨的同化、丙酮酸羧化还原成苹果酸等过程中起重要作用。
3.该途径中的一些中间产物是许多重要有机物质生物合成的原料,如Ru5P和R5P是合成核苷酸的原料。E4P和EMP 中的PEP可合成莽草酸,经莽草酸途径可合成芳香族氨基酸,还可合成与植物生长、抗病性有关的生长素、木质素、绿原酸、咖啡酸等。
4.该途径分子重组阶段形成的丙糖、丁糖、戊糖、己糖和庚糖的磷酸酯及酶类与卡尔文循环的中间产物和酶相同,因而戊糖磷酸途径和光合作用可以联系起来。
5.PPP在许多植物中普遍存在,特别是在植物感病、受伤、干旱时,该途径可占全部呼吸的50%以上。由于该途径和EMP-TCAC途径的酶系统不同,因此当EMP-TCAC途径受阻时,PPP则可替代正常的有氧呼吸。在糖的有氧降解中,EMP-TCAC途径与PPP所占的比例,随植物的种类、器官、年龄和环境而发生变化,这也体现了植物呼吸代谢的多样性。
五、乙醛酸循环
植物细胞内脂肪酸氧化分解为乙酰CoA之后,在乙醛酸体(glyoxysome)内生成琥珀酸、乙醛酸和苹果酸;此琥珀酸可用于糖的合成,该过程称为乙醛酸循环(glyoxylic acid cycle,GAC)。动物和人类细胞中没有乙醛酸体,无法将脂肪酸转变为糖。植物和微生物有乙醛酸体。油料植物种子(花生、油菜、棉籽等)萌发时存在着能够将脂肪转化为糖的乙醛酸循环。水稻盾片中也分离出了乙醛酸循环中的两个关键酶——异柠檬酸裂解酶和苹果酸合成酶。
图5-8 乙醛酸循环
(一)乙醛酸循环的化学历程
脂肪酸经过β-氧化分解为乙酰CoA,在柠檬酸合成酶的作用下乙酰CoA与草酰乙酸缩合为柠檬酸,再经乌头酸酶催化形成异柠檬酸。随后,异柠檬酸裂解酶(isocitratelyase)将异柠檬酸分解为琥珀酸和乙醛酸。再在苹果酸合成酶(malate synthetase)催化下,乙醛酸与乙酰CoA结合生成苹果酸。苹果酸脱氢重新形成草酰乙酸,可以再与乙酰CoA缩合为柠檬酸,于是构成一个循环(图5-8)。其总结果是由2分子乙酰CoA生成1分子琥珀酸,反应方程式如下:2乙酰CoA+NAD+→琥珀酸+2CoA+NADH+H+(5-14)
琥珀酸由乙醛酸体转移到线粒体,在其中通过三羧酸循环的部分反应转变为延胡索酸、苹果酸,再生成草酰乙酸。然后,草酰乙酸继续进入TCA循环或者转移到细胞质,在磷酸烯醇式丙酮酸羧激酶(PEP carboxykinase)催化下脱羧生成磷酸烯醇式丙酮酸(PEP),PEP再通过糖酵解的逆转而转变为葡萄糖6磷酸并形成蔗糖。
油料种子在发芽过程中,细胞中出现许多乙醛酸体,贮藏脂肪首先水解为甘油和脂肪酸,然后脂肪酸在乙醛酸体内氧化分解为乙酰CoA,并通过乙醛酸循环转化为糖,直到种子中贮藏的脂肪耗尽为止,乙醛酸循环活性便随之消失。淀粉种子萌发时不发生乙醛酸循环。可见,乙醛酸循环是富含脂肪的油料种子所特有的一种呼吸代谢途径。
以后在研究蓖麻种子萌发时脂肪→糖类的转化过程中,对上述乙醛酸循环途径作了修改。一是乙醛酸与乙酰CoA 结合所形成的苹果酸不发生脱氢,而是直接进入细胞质逆着糖酵解途径转变为蔗糖。二是在乙醛酸体和线粒体之间有“苹果酸穿梭”发生(图5-9)。
图5-9 修改后的脂肪酸通过乙醛酸循环转化为蔗糖的途径
由图5-9可以看出,通过“苹果酸穿梭”和转氨基反应解决了乙醛酸体内NAD+的再生和OAA的不断补充,这对保证GAC的正常运转是至关重要的。
(二)乙醛酸循环的特点和生理意义
1.乙醛酸循环和三羧酸循环中存在着某些相同的酶类和中间产物。但是,它们是两条不同的代谢途径。乙醛酸循环是在乙醛酸体中进行的,是与脂肪转化为糖密切相关的反应过程。而三羧酸循环是在线粒体中完成的,是与糖的彻底氧化脱羧密切相关的反应过程。
2.油料植物种子发芽时把脂肪转化为碳水化合物是通过乙醛酸循环来实现的。这个过程依赖于线粒体、乙醛酸体及细胞质的协同作用。
六、乙醇酸氧化途径
图5-10 水稻根中乙醇酸途径
乙醇酸氧化途径(glycolic acid oxidation pathway)是发生在水稻根系中的一种糖降解途径(图5-10)。水稻根呼吸产生的部分乙酰CoA不进入TCA循环,而是形成乙酸,然后乙酸在乙醇酸氧化酶及其它酶类催化下依次形成乙醇酸、乙醛酸、草酸和甲酸及CO2,并且不断地形成H2O2。H2O2在过氧化氢酶催化下产生具有强氧化能力的新生态氧,并释放于根的周围,形成一层氧化圈,使水稻根系周围保持较高的氧化状态,以氧化各种还原性物质(如H2S、Fe2+等),抑制土壤中还原性物质对水稻根的毒害,从而保证根系旺盛的生理机能,使稻株正常生长。
糖酵解途径
糖酵解途径(glycolytic pathway)是指细胞在胞浆中分解葡萄糖生成丙酮酸(pyruvate)的过程,此过程中伴有少量ATP的生成.在缺氧条件下丙酮酸被还原为乳酸(lactate)称为糖酵解.有氧条件下丙酮酸可进一步氧化分解生成乙酰CoA进入三羧酸循环,生成CO2和H2O. 葡萄糖不能直接扩散进入细胞内,其通过两种方式转运入细胞:一种是在前一节提到的与Na+共转运方式,它是一个耗能逆浓度梯度转运,主要发生在小肠粘膜细胞、肾小管上皮细胞等部位;另一种方式是通过细胞膜上特定转运载体将葡萄糖转运入细胞内(图4-1),它是一个不耗能顺浓度梯度的转运过程.目前已知转运载体有5种,其具有组织特异性如转运载体-1(GLUT-1)主要存在于红细胞,而转运载体-4(GLUT-4)主要存在于脂肪组织和肌肉组织. 糖酵解过程 糖酵解分为两个阶段共10个反应,每个分子葡萄糖经第一阶段共5个反应,消耗2个分子ATP为耗能过程,第二阶段5个反应生成4个分子ATP为释能过程. 1.第一阶段 (1)葡萄糖的磷酸化(phosphorylation of glucose) 进入细胞内的葡萄糖首先在第6位碳上被磷酸化生成6-磷酸葡萄糖(glucose 6 phophate,G-6-P),磷酸根由ATP供给,这一过程不仅活化了葡萄糖,有利于它进一步参与合成与分解代谢,同时还能使进入细胞的葡萄糖不再逸出细胞.催化此反应的酶是己糖激酶(hexokinase,HK).己糖激酶催化的反应不可逆,反应需要消耗能量
ATP,Mg2+是反应的激活剂,它能催化葡萄糖、甘露糖、氨基葡萄糖、果糖进行不可逆的磷酸化反应,生成相应的6-磷酸酯,6-磷酸葡萄糖是HK的反馈抑制物,此酶是糖氧化反应过程的限速酶(rate limiting enzyme)或称关键酶(key enzyme)它有同工酶Ⅰ-Ⅳ型,Ⅰ、Ⅱ、Ⅲ型主要存在于肝外组织,其对葡萄糖Km值为10-5~10-6M Ⅳ型主要存在于肝脏,特称葡萄糖激酶(glucokinase,GK),对葡萄糖的Km值1~10-2M,正常血糖浓度为5mmol/L,当血糖浓度升高时,GK 活性增加,葡萄糖和胰岛素能诱导肝脏合成GK,GK能催化葡萄糖、甘露糖生成其6-磷酸酯,6-磷酸葡萄糖对此酶无抑制作用. (2)6-磷酸葡萄糖的异构反应(isomerization of glucose-6-phosphate) 这是由磷酸己糖异构酶(phosphohexose isomerase)催化6-磷酸葡萄糖(醛糖aldose sugar)转变为6-磷酸果糖(fructose-6-phosphate,F-6-P)的过程,此反应是可逆的. (3)6-磷酸果糖的磷酸化(phosphorylation of fructose-6-phosphate) 此反应是6磷酸果糖第一位上的C进一步磷酸化生成1,6-二磷酸果糖,磷酸根由ATP供给,催化此反应的酶是磷酸果糖激酶1(phosphofructokinase l,PFK1). PFK1催化的反应是不可逆反应,它是糖的有氧氧化过程中最重要的限速酶,它也是变构酶,柠檬酸、ATP等是变构抑制剂,ADP、AMP、Pi、1,6-二磷酸果糖等是变构激活剂,胰岛素可诱导它的生成. (4)1.6 二磷酸果糖裂解反应(cleavage of fructose 1,6 di/bis phosphate)
三羧酸循环过程
三羧酸循环过程 乙酰-CoA进入由一连串反应构成的循环体系,被氧化生成H?O和CO?。由于这个循环反应开始于乙酰CoA与草酰乙酸(oxaloaceticacid)缩合生成的含有三 个羧基的柠檬酸,因此称之为三羧酸循环或柠檬酸循环(citratecycle)。在三羧酸循环中,柠檬酸合成酶催化的反应是关键步骤,草酰乙酸的供应有利于循环顺利进行。其详细过程如下:1、乙酰-CoA进入三羧酸循环乙酰CoA具有硫酯键,乙酰基有足够能量与草酰乙酸的羧基进行醛醇型缩合。首先柠檬酸合酶的组氨酸残基作为碱基与乙酰-CoA作用,使乙酰-CoA的甲基上失去一个h+,生成的碳阴离子对草酰乙酸的羰基碳进行亲核攻击,生成柠檬酰-CoA中间体,然后高能硫酯键水解放出游离的柠檬酸,使反应不可逆地向右进行。该反应由柠檬酸合成酶(citratesynthase)催化,是很强的放能反应。由草酰乙酸和乙酰-CoA 合成柠檬酸是三羧酸循环的重要调节点,柠檬酸合成酶是一个变构酶,ATP是柠檬酸合成酶的变构抑制剂,此外,α-酮戊二酸、NADH能变构抑制其活性,长链脂酰-CoA也可抑制它的活性,AMP可对抗ATP的抑制而起激活作用。2、异柠檬酸形成柠檬酸的叔醇基不易氧化,转变成异柠檬酸而使叔醇变成仲醇,就易于氧化,此反应由顺乌头酸酶催化,为一 可逆反应。3、第一次氧化脱羧在异柠檬酸脱氢酶作用下,异柠檬酸的仲醇氧化成羰基,生成草酰琥珀酸(oxalosuccinicacid)的中间产物,后者在同一酶表面,快速脱羧生成α-酮戊二酸(α-ketoglutarate)、NADH和CO2,此反应为β-氧化脱羧,此酶需要镁离子作为激活剂。此反应是不可逆的,是三羧酸循环中的限速步骤,ADP是异柠檬酸脱氢酶的激活剂,而ATP,NADH是此酶的抑制剂。4、第二次氧化脱羧在α-酮戊二酸脱氢酶系作用下,α-酮戊二酸氧化脱羧生成琥珀酰-CoA、NADH·H+和CO?,反应过程完全类似于丙酮酸脱氢酶系催化的氧化脱羧,属于α?氧化脱羧,氧化产生的能量中一部分储存于琥珀酰coa的高能硫酯键中。α-酮戊二酸脱氢酶系也由三个酶(α-酮戊二酸脱羧酶、硫辛酸琥珀酰基转移酶、二氢硫辛酸脱氢酶)和五个辅酶(tpp、硫辛酸、hscoa、NAD+、FAD)组成。此反应也是不可逆的。α-酮戊二酸脱氢酶复合体受ATP、GTP、NADH和琥珀酰-CoA抑制,但其不受磷酸化/去磷酸化的调控。5、底物磷酸化生成ATP 在琥珀酸硫激酶(succinatethiokinase)的作用下,琥珀酰-CoA的硫酯键水解,释放的自由能用于合成gtp,在细菌 和高等生物可直接生成ATP,在哺乳动物中,先生成GTP,再生成ATP,此时,琥珀酰-CoA生成琥珀酸和辅酶A。6、琥珀酸脱氢琥珀酸脱氢酶(succinatedehydrogenase)催化琥珀酸氧化成为延胡索酸。该酶结合在线粒体内膜上,而其他三羧酸循环的酶则都是存在线粒体基质中的,这酶含有铁硫中心和共价结合的fad,来自琥珀酸的电子通过fad和铁硫中心,然后进入电子传递链到O?,丙二酸是琥珀酸的类似物,是琥珀酸脱氢酶强有力的竞争性抑制物,所以可以阻断三羧酸循环。7、延胡索酸的水化延胡索酸酶仅对延胡索
糖酵解反应过程
有氧氧化的步骤简介 有氧氧化包括三个大的阶段,分别为糖的酵解、乙酰COA的 形成和三羧酸循环。 糖酵解反应过程 步骤名称底物酶产物能量 1 葡萄糖磷 酸化葡萄糖己糖激酶 HK 6-磷酸 葡萄糖 消耗ATP 一个 2 6-磷酸葡 萄糖异构6-磷酸葡 萄糖 葡萄糖己 糖异构酶 6-磷酸果 糖 3 6-磷酸果 糖磷酸化6-磷酸果 糖 磷酸果糖 激酶1 (PFK1) 1,6-二磷 酸果糖 消耗ATP 一个 4 磷酸丙糖 的生成1,6-二磷 酸果糖 缩醛酶磷酸二羟 丙酮,3- 磷酸甘油 醛 5 丙糖的转 化磷酸二羟 丙酮 磷酸丙糖 异构酶 3-磷酸甘 油醛 6 3-磷酸甘 油醛氧化 脱氢3-磷酸甘 油醛 3-磷酸甘 油脱氢酶 1,3-二磷 酸甘油酸 7 底物水平1,3-二磷磷酸甘油3-磷酸甘产生两分
磷酸化酸甘油酸酸激酶油酸子ATP 8 3-磷酸甘 油酸异构3-磷酸甘 油醛 磷酸甘油 酸变位酶 2-磷酸甘 油酸 9 2-磷酸甘 油酸烯醇 化2-磷酸甘 油酸 烯醇化酶磷酸烯醇 式甘油酸 10 底物水平 磷酸化磷酸烯醇 式甘油酸 丙酮酸激 酶 丙酮酸产生两分 子ATP 糖酵解过程简述 糖酵解在胞浆内进行,分为两阶段,第一阶段为3-磷酸甘油醛的生成,第二阶段为丙酮酸的生成。 第一阶段包括五个步骤 第一步为葡萄糖的磷酸化,葡萄糖在己糖激酶的催化下,消耗一个ATP,在6号c原子上挂上一个磷酸基,生成6-磷酸葡萄糖。第二步为6-磷酸葡萄糖的异构,在葡糖糖异构酶的催化下,6-磷酸葡萄糖异构为6-磷酸果糖。 第三步为6-磷酸果糖的磷酸化,在6-磷酸果糖激酶1的催化下,消耗一分子ATP,生成1,6-二磷酸果糖。 第四步为磷酸丙糖的生成,1,6-二磷酸果糖在缩醛酶的催化下生成一分子磷酸二羟丙酮和一分子3-磷酸甘油醛。 第五步为磷酸二羟丙酮的异构,磷酸二羟丙酮在丙糖异构酶的作用下生成3-磷酸甘油醛。
生物化学原理- 糖酵解
第十五章糖酵解 本章主线: 糖酵解 丙酮酸代谢命运 (乙醇发酵乳酸发酵) 糖酵解调控 巴斯德效应 3种单糖代谢 (果糖、半乳糖、甘露糖) 一、糖酵解 糖酵解概述: ●位置:细胞质 ●生物种类:动物、植物以及微生物共有 ●作用:葡萄糖分解产生能量 ●总反应:葡萄糖+2ADP+2 NAD++2Pi →2 丙酮酸+2ATP+2NADH+2H++2H2O 具体过程: 第一阶段(投入A TP阶段): 1分子葡萄糖转换为2分子甘油醛-3-磷酸;投入2分子ATP。 ○1 反应式:葡萄糖+ ATP→葡萄糖-6-磷酸+ADP 酶:己糖激酶(需Mg2+参与) 是否可逆:否 说明: ●保糖机制——磷酸化的葡萄糖被限制在细胞内,磷酸化的糖带有负电荷的磷酰基,可防 止糖分子再次通过质膜。(应用:解释输液时不直接输葡萄糖-6-磷酸的原因) ●己糖激酶以六碳糖为底物,专一性不强。 ●同功酶——葡萄糖激酶,是诱导酶。葡萄糖浓度高时才起作用。 ○2 反应式:葡萄糖-6-磷酸→果糖-6-磷酸 酶:葡萄糖-6-磷酸异构酶 是否可逆:是 说明:
●是一个醛糖-酮糖转换的同分异构化反应(开链?异构?环化) ●葡萄糖-6-磷酸异构酶表现出绝对的立体专一性 ●产物为α-D-呋喃果糖-6-磷酸 ○3 反应式:果糖-6-磷酸+ATP→果糖-1,6-二磷酸+ADP 酶:磷酸果糖激酶-I 是否可逆:否 说明: ●磷酸果糖激酶-I的底物是β-D-果糖-6-磷酸与其α异头物在水溶液中处于非酶催化的快 速平衡中。 ●是大多数细胞糖酵解中的主要调节步骤。 ○4 反应式:果糖-1,6-二磷酸→磷酸二羟丙酮+甘油醛-3-磷酸 酶:醛缩酶 是否可逆:是 说明: ●平衡有利于逆反应方向,但在生理条件下,甘油醛-3-磷酸不断地转化成丙酮酸,大大 地降低了甘油醛-3-磷酸的浓度,从而驱动反应向裂解方向进行。 ●注意断键位置:C3-C4 ○5 反应式:磷酸二羟丙酮→甘油醛-3-磷酸 酶:丙糖磷酸异构酶 是否可逆:是 说明: ●葡萄糖分子中的C-4和C-3 →甘油醛-3-磷酸的C-1; 葡萄糖分子中的C-5和C-2 →甘油醛-3-磷酸的C-2; 葡萄糖分子中的C-6和C-1 →甘油醛-3-磷酸的C-3。 ●缺少丙糖磷酸异构酶,将只有一半丙糖磷酸酵解,磷酸二羟丙酮堆积。 第二阶段(产出A TP阶段):此阶段各物质的量均加倍 2分子甘油醛-3-磷酸转换为2分子丙酮酸;产出4分子ATP ○6 反应式:甘油醛-3-磷酸+NAD++Pi→1,3-二磷酸甘油酸+NADH+H+ 酶:甘油醛-3-磷酸脱氢酶 是否可逆:是 说明: ●酵解中唯一一步氧化反应。是一步吸能反应,与第7步反应耦联有利于反应进行。 ●NAD+是甘油醛-3-磷酸脱氢酶的辅酶 ●1,3-二磷酸甘油酸中形成一个高能酸酐键。 ●无机砷酸(AsO43-)可取代无机磷酸作为甘油酸- 3-磷酸脱氢酶的底物,生成一个不稳
生物化学试题 糖酵解作用
第十九章糖酵解作用 一、是非判断题 1、ATP是果糖磷酸激酶的变构抑制剂。() 2、沿糖酵解途径简单逆行,可从丙酮酸等小分子前体物质合成葡萄糖。() 3、发酵可以在活细胞外进行。() 4、催化ATP分子中的磷酰基转移到受体上的酶称为激酶。() 5、糖酵解过程在有氧无氧条件下都能进行。() 6、在缺氧条件下,丙酮酸还原为乳酸的意义是使NAD+再生。() 二、填空题 1、1分子葡萄糖转化为2分子乳酸净生成______________分子ATP 2、糖酵解过程中有3个不可逆的酶促反应,这些酶是__________、____________ 和 _____________。 3、糖酵解抑制剂碘乙酸主要作用于___________酶。 4、糖酵解在细胞的_________中进行,该途径是将_________转变为_______,同时生成 ________和_______的一系列酶促反应。 5、乳酸脱氢酶在体内有5种同工酶,其中肌肉中的乳酸脱氢酶对__________ 亲和力特别高, 主要催化___________反应。 6、在糖酵解中提供高能磷酸基团,使ADP磷酸化成ATP的高能化合物是_______________ 和________________ 三、选择题 1、由己糖激酶催化的反应的逆反应所需要的酶是: A.果糖二磷酸酶B.葡萄糖-6-磷酸酶 C.磷酸果糖激酶D.磷酸化酶 2、正常情况下,肝获得能量的主要途径: A.葡萄糖进行糖酵解氧化B.脂肪酸氧化 C.葡萄糖的有氧氧化D.磷酸戊糖途径E.以上都是。 3、磷酸果糖激酶所催化的反应产物是: A.F-1-P B.F-6-P C.F-D-P D.G-6-P 4、醛缩酶的产物是: A.G-6-P B.F-6-P C.F-D-P D.1,3-二磷酸甘油酸 5、下面哪种酶在糖酵解和糖异生中都起作用: A.丙酮酸激酶B.丙酮酸羧化酶 C.3-磷酸甘油醛脱氢酶D.己糖激酶E.果糖1,6-二磷酸酯酶 6、糖酵解时哪一对代谢物提供P使ADP生成ATP: A.3-磷酸甘油醛及磷酸烯醇式丙酮酸 B.1,3-二磷酸甘油酸及磷酸烯醇式丙酮酸 C.1-磷酸葡萄糖及1,6-二磷酸果糖 D.6-磷酸葡萄糖及2-磷酸甘油酸
糖酵解特点
四、糖代谢概况 有氧 无氧 H 2O 及CO 2 乳酸 乳酸、氨基酸、甘油 糖原 核糖 + NADPH+H+ 磷酸戊糖途径 淀粉 消化与吸收 无氧分解(糖酵解) 糖酵解(glycolysis)是指葡萄糖在无氧条件下 分解生成乳酸并释放出能量的过程。 糖酵解的全部反应过程在胞液(cytoplasm)中进行,代谢的终产物为乳酸(lactate),一分子葡萄糖经无氧酵解可净生成两分子ATP 。 无氧酵解的反应过程可分为活化、裂解、放能 和还原四个阶段。
酸的生醇发酵及葡萄糖的无氧分解 葡萄糖EMP + NAD CH2OH CH3 乙醇 NADH+H+ NAD+ CO2 乳酸 COOH CH(O H) C H3 CHO CH3 COOH C==O CH3 丙酮酸 1.活化(a c t i v a t i o n)-己糖磷酸酯的生成 活化阶段是指葡萄糖经磷酸化和异构反应生成1,6-二磷酸果糖(FDP)的反应过程。该过程共由三步化学反应组成。 (一)糖酵解途径 葡萄糖磷酸化 磷酸葡萄糖(G-6-P) G-6-P异构为(F-6-P) F-6-P再磷酸化为( F-1,6-BP )......(1)......(2) (3)
ADP ATP ADP * (2 无氧酵解的活化阶段 第一阶段总结: 消耗ATP 不生成ATP 从葡萄糖开始→ 2分子ATP 从糖原开始→ 1分子ATP
.裂解(lysis)——磷酸丙糖的生 一分子F-1,6-BP 裂解为两分子可以互变的磷酸丙糖 (triose phosphate),包括两步反应: F-1,6-BP 裂解为3-磷酸甘油醛和磷酸二羟丙酮 磷酸二羟丙酮异构为3-磷酸甘油醛 (5) (4) 第二阶段总结: 1、一分子六碳糖分解为2分子能够互变的磷酸丙糖。 2、既不消耗ATP ,也不生成ATP 。 3.放能(r e l e a s i n g e n e r g y )——丙酮酸的生成 3-磷酸甘油醛经脱氢、磷酸化、脱水及放能等 反应生成丙酮酸,包括六步反应。 3- 磷酸甘油醛脱氢并磷酸化生成1,3- 二磷酸甘油酸 1,3-,将其交给ADP 生成ATP 3-磷酸甘油酸异构为2-磷酸甘油酸 (6) ......(7) (8)
糖酵解 三羧酸循环最全总结
在高等植物中存在着多条呼吸代谢的生化途径,这是植物在长期进化过程中,对多变环境条件适应的体现。在缺氧条件下进行酒精发酵和乳酸发酵,在有氧条件下进行三羧酸循环和戊糖磷酸途径,还有脂肪酸氧化分解的乙醛酸循环以及乙醇酸氧化途径等(图5-2)。 图5-2 植物体内主要呼吸代谢途径相互关系示意图 一、糖酵解 己糖在细胞质中分解成丙酮酸的过程,称为糖酵解(glycolysis)。整个糖酵解化学过程于1940年得到阐明。为纪念在研究这一途径中有突出贡献的三位生物化学家:G.Embden,O.Meyerhof和J.K.Parnas,又把糖酵解途径称为EmbdenMeyerhofParnas途径,简称EMP途径(EMP pathway)。糖酵解普遍存在于动物、植物、微生物的细胞中。 (一)糖酵解的化学历程 糖酵解途径(图5-3)可分为下列几个阶段:
图5-3糖酵解途径 1.己糖的活化(1~9)是糖酵解的起始阶段。己糖在己糖激酶作用下,消耗两个ATP逐步转化成果糖-1,6二磷酸(F-1,6-BP)。 如以淀粉作为底物,首先淀粉被降解为葡萄糖。淀粉降解涉及到多种酶的催化作用,其中,除淀粉磷酸化酶(starch phosphorylase)是一种葡萄糖基转移酶外,其余都是水解酶类,如α-淀粉酶(α-amylase)、β-淀粉酶(β-amylase)、脱支酶(debranching enzyme)、麦芽糖酶(maltase)等。 2.己糖裂解(10~11)即F-1,6-BP在醛缩酶作用下形成甘油醛-3-磷酸和二羟丙酮磷酸,后者在异构酶(isomerase)作用下可变为甘油醛-3-磷酸。 3.丙糖氧化(12~16)甘油醛-3-磷酸氧化脱氢形成磷酸甘油酸,产生1个ATP和1个NADH,同时释放能量。然后,磷酸甘油酸经脱水、脱磷酸形成丙酮酸,并产生1个ATP,这一过程分步完成,有烯醇化酶和丙酮酸激酶参与反应。
糖酵解的过程
糖酵解的過程EMP途徑分為兩個階段,第一 個階段是磷酸丙糖的生成過 程(耗能過程),第二和階段是丙酮酸生成過程(產能過程)。 下面讓我們來慢慢分解反應過程 第一階段第一步 △磷酸化:G→G6P Extracellular fluid:胞外液 Cytoplasm:細胞質 Glucose:葡萄糖 Phosphorylation:磷酸化作用 Plasma:等離子;血漿 Membrane:膜;薄膜第一階段 第二階段 己糖激酶 EMP 途 徑 中 第 一 個 限 速 酶激酶:一类从高能供体分子(如ATP)转移磷酸基团到特定靶分子(底物)的酶;这一过程谓之磷酸化。 已糖激酶:催化从ATP转移磷酸基团至各种六碳糖上去的酶。 激酶都需要Mg2+作为辅助因子。 首先我們來看一下糖酵解的第一階段
第一階段第二步△G6P F6P 第一階段第三步 ③磷酸化:F6P → FDP 磷酸葡萄糖异构酶 PFK是第二个限速酶,也是 EMP途径的关键酶,其活性 大小控制着整个途径的进 程。 磷酸果糖激酶是一种别构 酶,是糖酵解三个限速酶中 催化效率最低的酶,因此被 认为是糖酵解作用最重要 的限速酶。
第一階段第四步 ④裂解 (FBP → DHAP + G3P) 第一階段第⑤步 ⑤异构化(DHAP → G3P) 1,6-二磷酸果糖 2×3-磷酸甘油醛
第二階段第六步 ⑥氧化(G3P → 1,3-BPG) 第二階段第七步 ⑦转化(1,3-BPG → 3PG) 再來,我們來看糖 酵解的第二階段。 高能磷酸鍵 3-磷酸甘 ◎EMP第一次产生高能磷酸键; ◎EMP中唯一的脱氢反应,并产生了还原剂NADH。 ◎该酶是巯基酶,所以它可被碘乙酸不可逆地抑制,所以碘乙酸能抑制糖酵解。 ◎底物水平磷酸化:直接利用代谢 中间物氧化释放的能量产生ATP的 磷酸化类型。
糖酵解习题
糖代谢习题 1、写出在糖酵解反应过程中 A.在标准条件下耗能的反应 B.在标准条件下产能的反应 C.消耗ATP的反应 D.产生ATP的反应 E.在红细胞中处于或接近平衡的反应。 2、经过糖酵解途径后,如果丙酮酸分子中的某个碳原子被同 位素14C标记的话,请问这个碳原子在葡萄糖分子碳骨架中的哪个位置? 3、在红细胞中进行的糖酵解反应,如果下列物质浓度突然上 升会出现什么现象? A.ATP B. AMP C. 1,6-二磷酸果糖 D. 2,6-二磷酸果糖 E. 柠檬酸 F. 6-磷酸葡萄糖。 4、 1.3-二磷酸甘油酸水解生成3-磷酸甘油酸和无机磷酸(Pi) 的标准自由能(?G0’)是-49.6 kj/mol: 1.3-BPG + H20 →3-PG + Pi ATP水解的标准自由能是-30.5 kj/mol: ATP + H20 →ADP + Pi 1). 磷酸甘油酸激酶所催化的反应得标准自由能是多少? ADP + 1,3-BPG →ATP + 3-PG 2). 该反应的平衡常数是多少?
3). 如果红细胞中[1,3-BPG]和[3-PG]的浓度是1 μM,和120 μ M,假设磷酸甘油酸激酶催化的反应在平衡状态,请问[ATP]/[ADP] 是多少? 5、若进行柠檬酸循环的细胞中包含有磷酸烯醇式丙酮酸羧激酶,那么怎样的过程能使1分子的α-酮戊二酸氧化成5分子的CO2? 6、每进行一次三羧酸循环,1分子的乙酰CoA被氧化,即产生2分子的CO2。那么乙酰CoA的碳原子在其第一轮的循环中会被转变成CO2吗? 7、柠檬酸是对称分子,在柠檬酸循环的第二步中,乌头酸酶催化了半个柠檬酸分子脱水,却不催化相同的另一半脱水。这如何解释? 8、一系列的反应中,细胞需要NADPH的量远远超过5-p- 核糖的量。那么(a)细胞是如何获得NADPH的?(b) 过量的5-p-核糖有何去路? 9、丙二酸是琥珀酸脱氢酶催化反应的的一种竞争性抑制 剂。解释为什么增高草酰乙酸的浓度能够克服丙二酸的抑 制作用。假定这一反应发生在肝脏制剂中。 10、①丙氨酸降解产生丙酮酸,亮氨酸降解产生乙酰CoA。 这两种氨基酸的降解能为柠檬酸循环的中间物库作出补充 吗?②储存在动物脂肪组织的三酰甘油是能量的重要来 源。脂肪酸降解产生乙酰CoA,后者能激活丙酮酸羧化酶。
糖酵解过程详解
葡萄糖糖酵解详解 作者为了大家的方便,在网上搜集了资料,请交流,请提意见! 1,名称解析:在供氧不足时,体内组织细胞中的葡萄糖或糖元,分解为乳酸的过程称为无氧分解,由于此过程与与酵母菌使糖生醇发酵的过程基本相似,故称为糖酵解。 2,代谢位置:糖酵解是在细胞液中进行的。 3,过程可以分为两个阶段来理解: 第一阶段叫活化裂解阶段:由葡萄糖或糖元变成两分子磷酸丙糖(3-磷酸甘油醛和磷酸二羟丙酮),下面分别叙述: ○1如下图所示,为第一阶段的第○1小段。这一小段分两种情况:一个是从葡萄糖开始,一个是从糖元开始。 上图就表示从葡萄糖开始,葡萄糖首先在磷酸化酶催化下进行磷酸解,由ATP提供磷酸基生成6-磷酸葡萄糖,ATP本身变成ADP。大家注意代谢反应方程式的写法就是上面这个简化的表示式,相当于我们通常使用的下面的意思: 葡萄糖已糖激酶磷酸葡萄糖+ADP+H2O, 在这一阶段请注意: ▲从能量的角度来看,就消耗了一个ATP。但如果是从糖元开始,则因糖元在磷酸化酶催化下进行磷酸解是已变成了1-磷酸葡萄糖,下一步在变化酶作用下变成6-磷酸葡萄糖时就不消耗能量了,所以从糖元开始的糖酵解就少消耗这个ATP了。或者说因为糖原缩合时已经挂上了一分子磷酸,糖原一水解就是6磷酸葡萄糖,所以葡萄糖就不用再磷酸化了,就少消耗了一个atp。 ▲这阶段的已糖激酶是限速酶,决定反应的速度。 下面这图表示催化剂已糖酶的催化过程是把已糖酶把葡萄糖结合在一起形成1-磷酸葡萄糖(和6-磷酸葡萄糖是异构体)。 ○2第二小阶段是6-磷酸葡萄糖在已糖异构化酶催化下生成6-磷酸果糖,下面是这个反应的开链式和哈沃斯式的反应式:
糖酵解作用
糖酵解作用 填空题 1、1分子葡萄糖转化为2分子乳酸净生成 ______________分子ATP 2、糖酵解过程中有3个不可逆的酶促反应,这些酶是__________、____________ 和_____________。 3、糖酵解抑制剂碘乙酸主要作用于___________酶。 4、糖酵解在细胞的_________中进行,该途径是将 _________转变为_______,同时生成________和 _______的一系列酶促反应。 5、乳酸脱氢酶在体内有5种同工酶,其中肌肉中的乳酸脱氢酶对__________ 亲和力特别高,主要催化___________反应。 6、在糖酵解中提供高能磷酸基团,使ADP磷酸化成ATP的高能化合物是_______________ 和 ________________ 选择题 1、由己糖激酶催化的反应的逆反应所需要的酶是:A.果糖二磷酸酶B.葡萄糖-6-磷酸酶C.磷酸果糖激酶D.磷酸化酶
2、正常情况下,肝获得能量的主要途径: A.葡萄糖进行糖酵解氧化B.脂肪酸氧化C.葡萄糖的有氧氧化D.磷酸戊糖途 径E.以上都是。 3、磷酸果糖激酶所催化的反应产物是: A.F-1-P B.F-6-P C.F-D-P D.G-6-P 4、醛缩酶的产物是: A.G-6-P B.F-6-P C.F-D-P D.1,3-二磷酸甘油酸 5、下面哪种酶在糖酵解和糖异生中都起作用:A.丙酮酸激酶B.丙酮酸羧化酶 C.3-磷酸甘油醛脱氢酶D.己糖激酶 E.果糖1,6-二磷酸酯酶 6、糖酵解时哪一对代谢物提供P使ADP生成ATP:A.3-磷酸甘油醛及磷酸烯醇式丙酮酸 B.1,3-二磷酸甘油酸及磷酸烯醇式丙酮酸 C.1-磷酸葡萄糖及1,6-二磷酸果糖 D.6-磷酸葡萄糖及2-磷酸甘油酸 是非判断题 ()1、ATP是果糖磷酸激酶的变构抑制剂。
糖酵解三羧酸循环总结归纳
精心整理 在高等植物中存在着多条呼吸代谢的生化途径,这是植物在长期进化过程中,对多 变环境条件适应的体现。在缺氧条件下进行酒精发酵和乳酸发酵,在有氧条件下进行三 羧酸循环和戊糖磷酸途径,还有脂肪酸氧化分解的乙醛酸循环以及乙醇酸氧化途径等 (图5-2)。 图5-2植物体内主要呼吸代谢途径相互关系示意图 一、糖酵解 己糖在细胞质中分解成丙酮酸的过程,称为糖酵解(glycolysis)。整个糖酵解化学 1.糖酵解普遍存在于生物体中,是有氧呼吸和无氧呼吸途径的共同部分。 2.糖酵解的产物丙酮酸的化学性质十分活跃,可以通过各种代谢途径,生成不同的 物质(图5-4)。 图5-4丙酮酸在呼吸和物质转化中的作用 3.通过糖酵解,生物体可获得生命活动所需的部分能量。对于厌氧生物来说,糖酵 解是糖分解和获取能量的主要方式。 4.糖酵解途径中,除了由己糖激酶、磷酸果糖激酶、丙酮酸激酶等所催化的反应以 外,多数反应均可逆转,这就为糖异生作用提供了基本途径。 二、发酵作用 生物体中重要的发酵作用有酒精发酵和乳酸发酵。在酒精发酵(alcoholfermentation) 过程中,糖类经过糖酵解生成丙酮酸。然后,丙酮酸先在丙酮酸脱羧酶
(pyruvicaciddecarboxylase)作用下脱羧生成乙醛。 CH3COCOOH→CO2+CH3CHO(5-5) 乙醛再在乙醇脱氢酶(alcoholdehydrogenase)的作用下,被还原为乙醇。 CH3CHO+NADH+H+→CH3CH2OH+NAD+(5-6) 在缺少丙酮酸脱羧酶而含有乳酸脱氢酶(lacticaciddehydrogenase)的组织里,丙酮酸便被NADH还原为乳酸,即乳酸发酵(lactatefermentation)。 CH3COCOOH+NADH+H+→CH3CHOHCOOH+NAD+(5-7) 在无氧条件下,通过酒精发酵或乳酸发酵,实现了NAD+的再生,这就使糖酵解得以继续进行。 无氧呼吸过程中形成乙醇或乳酸所需的NADH+H+,一般来自于糖酵解。因此,当植物进行无氧呼吸时,糖酵解过程中形成的2分子NADH+H+就会被消耗掉(图5-5), (thiaminepyrophosphate,TPP)、辅酶A(coenzymeA)、硫辛酸(lipoicacid)、 FAD(flavinadeninedinucleotide)、NAD+(nicotinamideadeninedinucleotide)和Mg2+。 图5-6三羧酸循环的反应过程 上述反应中从底物上脱下的氢是经FAD→FADH2传到NAD+再生成NADH+H+。 2.反应(2)乙酰CoA在柠檬酸合成酶催化下与草酰乙酸缩合为柠檬酸,并释放CoASH,此反应为放能反应(△G°,=-32.22kJ·mol-1)。 3.反应(3)由顺乌头酸酶催化柠檬酸脱水生成顺乌头酸,然后加水生成异柠檬酸。 4.反应(4)在异柠檬酸脱氢酶催化下,异柠檬酸脱氢生成NADH,其中间产物草酰琥珀酸是一个不稳定的β-酮酸,与酶结合即脱羧形成α-酮戊二酸。 5.反应(5)α酮戊二酸在α酮戊二酸脱氢酶复合体催化下形成琥珀酰辅酶A和NADH,并释放CO2。
糖酵解的生理意义
糖酵解的生理意义:产生ATP;提供生物合成的原料;糖酵解与肿瘤:缺氧与缺氧诱导的转录因子HIF-1,DNA上的缺氧应答元件;参与糖酵解途径的一些酶的兼职功能。C6H12O6+2Pi +2ADP+2NAD→2CH3COCOOH+2ATP + 2H2O+2NADH +2H;;TCA 循环乙酰-CoA+3NAD+FAD+GDP+Pi+2H2O→2CO2+3NADH+FADH2+GTP+2H++CoA;;TCA循环的功能:产生更多的ATP;提供多种生物分子合成的原料;是糖、氨基酸和脂肪酸最后的共同分解途径;某些代谢中间物作为其他代谢途径的别构效应物;产生CO2;乙醛酸循与三羧酸循环区别:在每一轮循环中,有两分子乙酰-CoA进入,净生一分子琥珀酸;只产生NADH,但不产生FADH2;无底物水平磷酸化反应,因此不产生ATP;不生成CO2,无碳单位的损失;具有此途径的生物能够以乙酸作为唯一碳源。磷酸戊糖途径的功能:与NADPH 有关的功能1. 提供生物合成的还原剂NADPH2. 解毒——细胞色素P450单加氧酶解毒系统需要NADPH参与对毒物的羟基化反应。3. 免疫4. 维持红细胞膜的完整5. 间接进入呼吸链;提供核苷酸及其衍生物合成的前体5-磷酸核糖;提供芳香族氨基酸和维生素B6的合成需要的赤藓糖。糖异生的生理功能在饥饿或碳水化合物摄入不足的情况下,可以补充血糖,维持血糖浓度的稳定,为那些特别依赖葡萄糖氧化放能的细胞和组织提供燃料;植物和某些微生物使用乙酸作为糖异生的前体,使得它们能以乙酸作为唯一碳源;减轻或消除代谢性酸中毒。机体使用糖原作为能量储备的理由首先,糖原动员起来更为容易,因为它是高度分支的分子,糖原的磷酸解反应可以在各非还原端同时展开;其次,糖原的分解以及后面的糖酵解既可以在有氧又可以在无氧的条件下进行;动物体内偶数脂肪酸无法转化为葡萄糖,当饥饿的时候,肝糖原可迅速分解并转化为血糖,为脑组织等提供燃料。糖原合成 1.需要活化的葡萄糖单位——UDPG(动物和酵母)细菌(ADPG) 2.需要引物——一是没有完全降解的糖原分子;二是糖原素或糖原蛋白 3.糖原合成方向:从还原端向非还原端进行 4.分支需要分支酶。β-氧化的功能:产生ATP,其产生ATP的效率要高于葡萄糖。产生大量的H2O。这对于某些生活在干燥缺水环境的生物十分重要,像骆驼已将β-氧化作为获取水的一种特殊手段。脂肪酸合成的反应历程:1.引发反应:作为引物的乙酰基从乙酰辅酶A转移到脂肪酸合酶的一个亚基上。2.活化的“二碳单位”的装载。3.缩合是一步碳链延伸的反应。 4.还原以KR催化NADPH为还原剂的氧化还原反应 5.脱水 6.再还原 7.软脂酸的释放8乙酰-CoA+14NADPH+7H++7ATP→软脂酸+14NADP++8HSCoA+6H2O+7ADP+7Pi联合脱氨反应:由转氨酶和谷氨酸脱氢酶组合在一起的脱氨基反应。作用次序:先在转氨酶的催化下,一种氨基酸的氨基被转移到α-酮戊二酸的羰基上形成Glu,然后Glu在谷氨酸脱氢酶的催化下发生氧化脱氨反应,产生α-酮戊二酸、NH4+和NAD(P)H。氨基酸“X”+α-酮戊二酸→谷氨酸+α-酮酸“X”→α-酮戊二酸+NH4++NAD(P)H。从头合成:从最简单的小分子,如CO2和氨基酸等开始,经过多步反应,消耗更多的能量,最后生成核苷酸的过程。补救途径:指核苷酸降解的中间产物(包括核苷和碱基)被循环利用,重新转变成核苷酸的过程。嘌呤环上各原子来源:1N天冬氨酸,2C8C N10-甲酰-四氢叶酸,3N9N 谷氨酰胺,4C5C7N甘氨酸,6CCO2嘧啶核苷酸的从头合成前体:Gln、CO2、Asp等。与嘌呤核苷酸从头合成的区别:嘌呤核苷酸是先形成β-N-糖苷键,然后再逐步形成嘌呤环。而嘧啶核苷酸是先形成嘧啶环,然后再与PRPP形成β-N-糖苷键;嘌呤核苷酸从头合成所有的反应都是在细胞质基质内发生的,而嘧啶核苷酸从头合成的反应则发生在线粒体。DNA复制的一般特征①以原来的DNA两条链作为模板,四种dNTP为前体,还需要Mg2+②作为模板的DNA需要解链③半保留复制④需要引物——主要是RNA,少数是蛋白质⑤复制的方向始终是5′→3′⑥具有固定的起点⑦多为双向复制,少数为单向复制⑧半不连续性⑨具有高度的忠实性和进行性DNA聚合酶的全名是依赖于DNA的DNA聚合酶,就是以DNA为模板,催化DNA合成的聚合酶。该酶不能催化DNA的从头合成决定了DNA复制需要引物,而它只能从5'→3'催化聚合决定了DNA复制的单向性。DNA聚合酶Ⅰ具有5'→3'的DNA聚合酶活性还具有5'-外切酶和3'-外切酶活性。复制体:由DNA和多种蛋白质组装而成的催化DNA 复制的复合体。DNA 转录一般特征(与DNA复制的比较):与DNA复制的共同性质1.需要模板、解链和解除转录过程中形成的正超螺旋2.只能按照5′→3′的方向进行与DNA复制不同的性质1.不需要引物2.NTPs 代替dNTPs; UTP代谢dTTP3.缺乏校对活性4.发生在特定的区域(不是所有的DNA序列)5.对于一个特定的基因而言,只有一条链转录1.转录具有选择性和不对称性,只发生在DNA分子上的某些特定区域。2.以四种NTP--ATP、GTP、CTP和UTP为前体还需要Mg2+ 。3.转录需要模板、解链,但不需要引物。4.最先转录出来的核苷酸通常为嘌呤核苷酸。5.转录方向总是从5’→3’。6.转录也具有高度的忠实性,但是比DNA复制低。 7.有高度的进行性。8..转录受到严格的调控。细菌RNA聚合酶抑制剂:利福霉素和利链霉素启动子在转录起始点的上游存在着一些特殊的高度保守性的碱基序列。帽子结构功能:提高mRNA的稳定性;参与识别起始密码子的过程,提高mRNA的可翻译性;有助于mRNA通过核孔从细胞核运输到细胞质;提高剪接反应的效率。核糖体:核糖体是一种复杂的核糖核蛋白颗粒,由大小两个亚基组成,存在于原核细胞和真核细胞的细胞质中,以及真核细胞的线粒体和叶绿体中。核糖体的功能位点:1. A部位—氨酰tRNA结合部位,也称为受体部位;2. P 部位—肽酰tRNA结合部位;3. E部位—空载tRNA临时结合的部位;4. 肽酰转移酶活性部位——催化肽键形成的部位(大亚基);5. mRNA结合部位;6. 多肽链离开通道——正在延伸的多肽链离开核糖体的通道;7. 一些可溶性蛋白质因子(起始因子、延伸因子和终止因子)的结合部位。mRNA作为翻译的模板,至少含有一个ORF,即以起始密码子开始、以终止密码子结束的一段连续的核苷酸序列。tRNA:在翻译中,tRNA 是一种双功能接头分子,一头与氨基酸结合,一头通过反密码环的反密码子与mRNA结合,对mRNA进行解码。翻译的一般特征:1. mRNA、tRNA和核糖体起相同的作用2. 翻译的极性:阅读mRNA的方向都是从5′端→3′端,多肽链生长的方向总是从N-端→C-端。3. 遗传密码是三联体密码4. 正确的氨基酸的参入取决于mRNA上的密码子与tRNA上的反密码子之间的相互作用,与tRNA所携带的氨基酸无关5. 密码子与反密码子的相互识别遵守摆动规则6. 在核糖体上同源tRNA的识别是诱导契合的过程三联体密码性质:1.简并与兼职2.密码子的摆动性3.通用和例外4.不重叠,无标点5.方向性6.同一种氨基酸的不同密码子使用的频率不尽相同。因子结构与功能:IF2(GTP)-促进起始tRNA与核糖体小亚基结合;IF3-核糖体的解离和mRNA的结合;EF-G-结合核糖体和GTP,促进核糖体移位;RF1识别UAA或UAG,RF2识别UAA或UGA;RRF-翻译终止后,促进核糖体解体。操纵子模型认为:一些功能相关的结构基因成簇存在,构成所谓的多顺反子,它们的表达作为一个整体受到控制元件的调节。控制元件由启动子、操纵基因)和调节基因组成。调节基因编码调节蛋白,与操纵基因结合而调节结构基因的表达。
糖酵解的过程
糖酵解的過程 EMP 途徑分為兩個階段,第一個階段是磷酸丙糖的生成過程(耗能過程),第二和階段是丙酮酸生成過程(產能過程)。 下面讓我們來慢慢分解反應過程 第一階段 第一步 △一 磷酸化:G →G6P Extracellular fluid :胞外液 Cytoplasm :細胞質 Glucose :葡萄糖 Phosphorylation :磷酸化作用 Plasma :等離子;血漿 Membrane :膜;薄膜 第一階段 第二階段 己糖激酶 EMP 途 徑中第一個限速酶 激酶:一类从高能供体分子(如ATP )转移磷酸基团到特定靶分子(底物)的酶;这一过程谓之磷酸化。 已糖激酶:催化从ATP 转移磷酸基团至各种六碳糖上去的酶。 激酶都需要Mg2+作为辅助因子。 首先我們來看一下糖酵解的第一階段
第一階段第二步△二 G6P F6P 磷酸葡萄糖异构酶 第一階段第三步 ③磷酸化:F6P → FDP PFK是第二个限速酶,也是 EMP途径的关键酶,其活 性大小控制着整个途径的 进程。 磷酸果糖激酶是一种别构 酶,是糖酵解三个限速酶中 催化效率最低的酶,因此被 认为是糖酵解作用最重要 的限速酶。
第一階段 第四步 ④ 裂解 (FBP → DHAP + G3P ) 第一階段 第⑤步 ⑤ 异构化(DHAP → G3P ) 1,6-二磷酸果糖 2×3-磷酸甘油醛 ◎上述5步反应完成了糖酵解的准备阶段。 ◎包括两个磷酸化步骤,由六碳糖裂解为两分子三碳糖,最后都转变为3-磷酸甘油醛。 ◎在准备阶段中,并没有从中获得任何能量,与此相反,却消耗了两个ATP 分子。 ◎以下的5步反应包括氧化-还原反应、磷酸化反应。这些反应正是从3-磷酸甘油醛提取能量形成ATP 分子。
三羧酸循环
第23章三羧酸循环(生物化学下册p92) 3学时 学习重点: ◆熟悉柠檬酸循环途径中的各步酶促反应,以及各步反应酶的作用特点。 ◆会分析和计算酵解和柠檬酸循环中产生的能量,以及底物分子中标记碳的去向。 葡萄糖的有氧氧化包括四个阶段。 ①糖酵解产生丙酮酸(2丙酮酸、2ATP、2NADH) ②丙酮酸氧化脱羧生成乙酰CoA ③三羧酸循环(CO2、H2O、A TP、NADH) ④呼吸链氧化磷酸化(NADH-----ATP) 三羧酸循环:乙酰CoA经一系列的氧化、脱羧,最终生成CO2、H2O、并释放能量的过程,又称柠檬酸循环、Krebs循环。 原核生物:①~④阶段在胞质中 真核生物:①在胞质中,②~④在线粒体中 一、丙酮酸脱羧生成乙酰CoA 1、反应式: 2、丙酮酸脱氢酶系 丙酮酸脱氢酶系是一个十分庞大的多酶体系,位于线粒体膜上,电镜下可见。 E.coli丙酮酸脱氢酶复合体: 分子量:4.5×106,直径45nm,比核糖体稍大。 酶辅酶每个复合物亚基数 丙酮酸脱羧酶(E1)TPP 24 二氢硫辛酸转乙酰酶(E2)硫辛酸24 二氢硫辛酸脱氢酶(E3)FAD、NAD+12 此外,还需要CoA、Mg2+作为辅因子 这些肽链以非共价键结合在一起,在碱性条件下,复合体可以解离成相应的亚单位,在中性时又可以重组为复合体。所有丙酮酸氧化脱羧的中间物均紧密结合在复合体上,活性中间物可以从一个酶活性位置转到另一个酶活性位置,因此,多酶复合体有利于高效催化反应及调节酶在反应中的活性。 3、反应步骤 反应过程 (1)丙酮酸脱羧形成羟乙基-TPP
(2)二氢硫辛酸乙酰转移酶(E2)使羟乙基氧化成乙酰基 (3)E2将乙酰基转给CoA,生成乙酰-CoA (4)E3氧化E2上的还原型二氢硫辛酸 (5)E3还原NAD+生成NADH 4、丙酮酸脱氢酶系的活性调节 从丙酮酸到乙酰CoA是代谢途径的分支点,此反应体系受到严密的调节控制,此酶系受两种机制调节。 (1)可逆磷酸化的共价调节 丙酮酸脱氢酶激酶(E A)(可被ATP激活) 丙酮酸脱氢酶磷酸酶(E B) 磷酸化的丙酮酸脱氢酶(无活性) 去磷酸化的丙酮酸脱氢酶(有活性) (2)别构调节 ATP、CoA、NADH是别构抑制剂 ATP抑制E1 CoA抑制E2 NADH抑制E3 5、能量 1分子丙酮酸生成1分子乙酰CoA,产生1分子NADH(2.5A TP)。 二、三羧酸循环(TCA)的过程 TCA循环:每轮循环有2个C原子以乙酰CoA形式进入,有2个C原子完全氧化成CO2放出,分别发生4次氧化脱氢,共释放10A TP。 1、反应步骤 概述三羧酸循环(图,见书) (1)、乙酰CoA+草酰乙酸→柠檬酸 柠檬酸合酶,TCA中第一个调节酶:受ATP、NADH、琥珀酰CoA、和长链脂肪酰CoA 的抑制;受乙酰CoA、草酸乙酸激活。 柠檬酸合酶上的两个His残基起重要作用: 一个与草酰乙酸羰基氧原子作用,使其易受攻击;另一个促进乙酰CoA的甲基碳上的质子离开,形成烯醇离子,就可与草酰乙酸缩合成C-C键,生成柠檬酰CoA,后者使酶构象变化,使活性中心增加一个Asp残基,捕获水分子,以水解硫酯键,然后CoA和柠檬酸相继离开酶。 氟乙酰CoA可与草酰乙酸生成氟柠檬酸,抑制下一步反应的酶,据此,可以合成杀虫剂、灭鼠药。 氟乙酸本身无毒,氟柠檬酸是乌头酸酶专一的抑制剂,氟柠檬酸结合到乌头酸酶的活性部位上,并封闭之,使需氧能量代谢受毒害。它存在于某些有毒植物叶子中,是已知最能致死的简单分子之一。LD50为0.2mg/Kg体重,它比强烈的神经毒物二异丙基氟磷酸的LD50
糖酵解、TCA途径
糖酵解途径(EMP途径) 定义:葡萄糖经过一系列步骤降解成丙酮酸并生成ATP过程,被认为是微生物最古老原始的获能方式。指在O2不足情况下,葡萄糖或糖原分解为丙酮酸或乳酸,并伴随少量ATP生成。在细胞质中进行。 两个阶段: 一:活化阶段 a:葡萄糖磷酸化:活化葡萄糖,消耗1ATP,使葡萄糖和磷酸结合成葡萄糖-6-磷酸(己糖激酶) b:葡萄糖-6-磷酸重排成果糖-6-磷酸(葡萄糖磷酸异构酶) c:生成果糖-1、6-二磷酸(6-磷酸果糖激酶-1),消耗1ATP d:果糖-1、6-二磷酸断裂为3-磷酸甘油醛和磷酸二羟丙酮(醛缩酶)e:磷酸二羟丙酮很快转变为3-磷酸甘油醛。(丙糖磷酸异构酶)二:放能阶段 a:3-磷酸甘油醛氧化生成1、3-二磷酸甘油酸,释出2电子和1H+,生成NADH+ H+,且将能量转移至高能磷酸键中。 b:不稳定的1、3-二磷酸甘油酸失去高能磷酸键,生成3-磷酸甘油酸,能量转移至ATP中,生成1ATP(发生第一次底物水平磷酸化)c:3-磷酸甘油酸重排生成2-磷酸甘油酸 d:2-磷酸甘油酸脱水生成磷酸烯醇式丙酮酸 e:磷酸烯醇式丙酮酸将磷酸基团转移给ADP生成ATP,同时形成丙酮酸(发生第一次底物水平磷酸化)
附图:
总反应式: 一.糖无氧氧化反应(分为糖酵解途径和乳酸生成两个阶段)(一)糖酵解的反应过程(不是限速酶的反应均是可逆的) 1.葡萄糖磷酸化为6-磷酸葡萄糖 [1] 己糖激酶(hexokinase)催化,I-IV型,肝细胞中为IV型,又称葡萄糖激酶 区别:前者Km值小、特异性差。 意义:浓度较低时,肝细胞不能利用Glc。 [2]需要Mg++参与,消耗1分子ATP [3] 关键酶(限速酶):己糖激酶。 [4]反应不可逆,受激素调控。 [5]磷酸化后的葡萄糖不能透过细胞膜而逸出细胞。